Transpiration/Evaporation Ratio in Prunus Fremontii and Marianna 2624 over a 4-Day Period of Drought - Juniper Publishers
Juniper Publishers- Open Access Journal of Environmental Sciences & Natural Resources

Transpiration/Evaporation Ratio in Prunus Fremontii and Marianna 2624 over a 4-Day Period of Drought
Authored by Riccardo Lo Bianco
Abstract
Drought stress represents one of the major environmental limitations to crop production in a large portion of the Earth's surface. Natural genetic resources are one of the most powerful weapons against environmental stress. In such contest, the behaviour in terms of water loss of two species, P. fremontii (slower growing) and Marianna 2624 (faster growing), native to arid and humid regions respectively, was tested. Transpiration (T) and evapo transpiration (ET) rates and soil water content (SWC) were measured in the two species under controlled conditions. Evaporation rate from the soil surface was obtained by difference, and an average transpiration/evaporation ratio (T/E) was calculated to determine the relative importance of the two processes. Soil water content decreased linearly in both species over the 4-day period, however only Marianna 2624 ET decreased along with SWC. On the contrary, T and ET of both species correlated positively with temperature, indicating a more direct response at the leaf level. A much higher T/E in Marianna 2624 than in P. fremontii also indicated that water was lost mainly by T in the faster-growing species, while evaporation from the soil surface was the predominant process of water loss in the slower-growing species.
Keywords: Drought; Evapo Transpiration; Fast Growing; Slow Growing; Soil Water Content
Introduction
Only 7.6 % of the Earth's surface is suitable for agriculture; the remainder has one or more environmental limitations to crop production such as drought, saline soil, or extremes in temperature. On cultivated land, it is estimated that environmental stresses limit agricultural production to perhaps 25% of its potential level in the United States [1], and global climate changes are constantly reducing such percentage. Environmental stress tolerance is therefore a critical concern for agriculturists if they hope to increase food production as population increases. In particular, water loss from plant tissue (drought stress) may cause decreases in fruit quantity and quality. The status of the water- storage elements soil properties, vegetation, and topography determines the quantity of precipitation that may be retained and the extent to which the evaporative demand can be satisfied. In turn, the evaporative demand is a function of the atmospheric environment (air temperature, relative humidity, wind speed, and solar radiation [2]. Over the United States as a whole, it is estimated that about 70% of the annual precipitation is lost to evapo transpiration (ET, up to 90% in areas of the arid west) [3]. This fact proves the importance of adequately estimating the ET component of the hydrologic cycle in predicting on-farm irrigation water management and irrigation planning [4], especially if we consider that without ET there is no production [5]. The partitioning of the transpiration and evaporation components, as well as their ratio, have been primarily used to study vegetation distribution and water consumption across specific areas Herbs, [6]. Similarly, the transpiration to ET coefficients has been widely used for precise and efficient irrigation management [7]. In this contest, understanding the transpiration mechanisms of plants and the factors that control evaporation from the soil surface becomes a priority. Green plants have in fact many structures and control devices, which allow them to function efficiently even in rapidly changing environments. At root level, for example, it is to the advantage of the plant to expose a large surface to the soil and to sample as largely a volume of soil as possible. When the soil is wet the water enters all root surfaces including those that are old and suberized. Conversely, fine roots and root hairs seem to be more effective in fairly dry soil, possibly because of aeration problems [8]. At the leaf level, transpiration is controlled mainly by physiological and structural factors as follows:
T = gs * A/D
Where T is the transpiration rate, gs the stomatal conductance, A the stomatal aperture and D the depth of the stomatal pore [9]. Stomatal conductance responds to several factors such as light, CO2 concentration, vapor pressure deficit, leaf temperature, and soil water potential. This latter factor influences gs by a hormonal signal (abscissic acid) originating in the roots, a sort of biological switch when drought occurs [10]. The ratio A/D represents an index of the shape of the stomatal pore and can vary widely in different species, often depending upon the environmental conditions of native areas. A case in point is the sunken stomata. The guard cells of a sunken stomata are in a depression below the level of the leaf surface, and the walls of this depression have cuticle on their surfaces, so that, even when the stomata is open, gas or water vapor must diffuse through some distance. Therefore, water loss through sunken stomata does not increase with wind as much as it does through non-sunken stomata. Leaf hairs (same effect as sunken stomata), reduced leaf size (needle), wax cover on the leaf, etc. are other examples of structures that plants use to reduce water loss. Plants may also adapt genetically to long-term climate changes by changing life strategy [11,12]; this is the case, for example, of a slow growing species in an arid climate that maintains its turgor by reducing transpiration, but at the same time reduces CO2 assumption and therefore growth. On the other hand, evaporation from the soil surface results to be regulated by many factors related to the soil (type, texture, colour, organic matter content and type, capillary activity) and its water content. Another important factor affecting evaporation is the shading effect (leaf area index, canopy structure, etc.) due to the vegetation that indirectly affects the soil temperature [13]. For the above reasons, knowing the partitioning of water loss between plant and soil may be useful to discriminate plant behaviour under dry environments and select for more resilient genotypes. In this contest, the amount of water lost by transpiration versus the one lost by evaporation from the soil surface was tested in two species native to different climatic regions, over a 4-day period during which no water was supplied (from wet to dry soil conditions). It was hypothesized that a) the transpiration/evaporation ratio was greater in the faster-growing species; b) the ET rate of the faster- growing species decreased faster than that of the slower growing species from wet to dry soil conditions.
Materials and Methods
The experiment was conducted in a greenhouse where temperature was controlled, ranging from a minimum of 18 °C to a maximum of 32 °C, and the relative humidity was the same as outside. Two species with different characteristics were selected, the slower-growing Prunus fremontii native to south California that presents an average leaf dimension of 0.9-1.2 cm and is better adapted to arid climates; and the faster-growing Marianna 2624, a hybrid between Prunus cerasifera and Prunus munsoniana commonly used as rootstock in fruit crop production, with an average leaf size of 4-5 cm and better adapted to humid climates. Sixteen 1-year-old seedlings (eight from each species) with similar LAI and canopy structure were transplanted in 3-liter pots containing 100% pine bark as soil medium. Data of water lost only by transpiration (T) were collected by weighing plants where the pot was enclosed in white plastic bags tied up at the bottom of the stem. The plastic bags were kept open at night-time to allow root respiration. Data of total water lost by T and evaporation (E) were collected from control plants. Also in this case, the same white plastic bags were wrapped on the external surface of the pots to maintain the same root temperature. Measurements were made every day at noon and at sunset (8:30 p.m.) over a period of four days as Marianna seedlings started showing wilting symptoms. Plants were watered to field capacity only once at the onset of the experiment. The level of soil moisture was determined throughout the four days by subtracting the weight of the pot, dry pine bark and plant to the total weight. Standard error of the means and linear regression were used to analyze data.
Results and Discussion
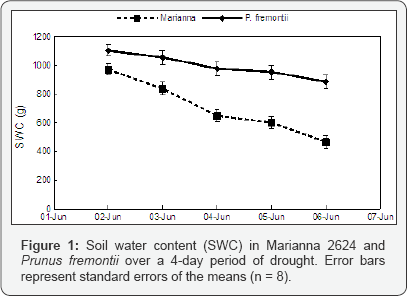
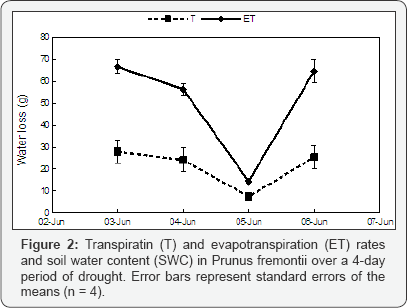
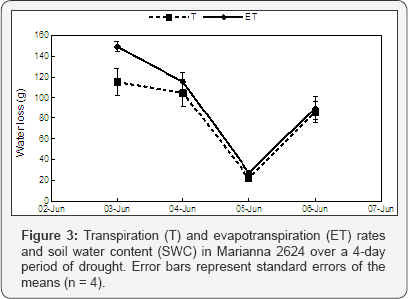
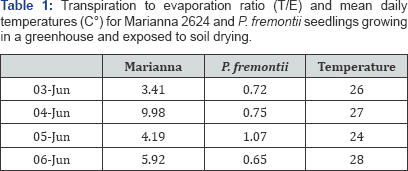
The soil water content (SWC) decreased significantly over the 4-day period in both P. fremontii and Marianna 2624 (P = 0.017 and P = 0.024 respectively, n = 16; (Figure 1). However, in P. fremontii neither ET nor T rates followed SWC (P = 0.560 and P = 0.522 respectively, n = 16) probably because the total loss of water from the soil (%) was relatively small over the 4-day period (Figure 2). A drop in ET as well as in T occurred only on June 5 when solar radiation and temperature were relatively low (rainy day, 24 °C). In fact, a positive correlation (P = 0.018, n = 16) was found between T and temperature indicating a more direct effect of temperature, and probably solar radiation, than SWC on plant transpiration. Conversely, in Marianna 2624, where a bigger amount of water was lost by ET than in P. fremontii, a significant decrease in ET (P = 0.018, n = 16), but not in T (P = 0.132, n = 16) is detectable as the soil dries out (Figure 3). This may indicate that SWC has a more direct effect on the evaporation from the soil surface than on plant transpiration at least at these levels of SWC. Similarly to P. fremontii, also in Marianna 2624, a positive correlation (P = 0.010, n = 16) between T and temperature confirmed the importance of temperature and solar radiation for gas exchange. The transpiration/evaporation ratio (T/E) was not influenced by soil moisture, and it showed higher values in Martianna 2624 than in P. fremontii (Table 1), indicating a predominant loss of water by transpiration in the faster-growing species. In particular, on June 4 (mostly sunny, 27 °C) T/E of Marianna 2624 reached its highest value and was over 10 folds higher than in P. fremontii (Table 1). This furthermore indicates a stronger metabolic response (photosynthesis) of the faster-growing species to temperature and solar radiation as compared to that of the slower-growing species. On the other hand, evaporation seemed to become the predominant way of water loss in systems with slower-growing species, where transpiration is relatively low. The latter surely represents a conservative strategy in terms of growth, but that may become of advantage to the plant in situations of prolonged and/ or severe moisture deficiency. In conclusion, the results obtained here confirm the hypothesis that different species can utilize water differently depending upon their defence strategy against environmental stresses. In particular, faster-growing species seem to respond more quickly to changes in temperature and solar radiation showing, however, lesser ability to tolerate drought stress than slower-growing species. Quantifying the importance of climatic factors such as solar radiation, air temperature, wind speed, etc., and evaluating the interaction between these factors and the genetic differences among species will better explain the mechanisms underlying the biological response to different climates.
For more articles in Open Access Journal of Environmental Sciences & Natural Resources please click on: https://juniperpublishers.com/ijesnr/index.php



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Comments
Post a Comment